La Moelle épinière (3) Anatomie fonctionnelle
5.4.2.2.-Les voies descendantes (voies motrices)
5.4.2.2.1.-Organisation générale S.64

Il existe deux catégories des voies motrices :
Les unes sont directes et monosynaptiques et constituent la voie cortico-spinale directe, présente dans le faisceau pyramidal. Elle est constituée par une chaîne de deux neurones.
Le premier neurone est le neurone central, au trajet strictement cortico-spinal. Sa lésion pathologique, quel qu’en soit le niveau (cerveau, tronc cérébral, moelle épinière), se traduit par une paralysie centrale, avec spasticité.
Le deuxième neurone est le motoneurone de la corne ventrale ou motoneurone périphérique dont l’axone forme les fibres nerveuses motrices des racines spinales et des nerfs périphériques. Sa lésion pathologique se traduit par une paralysie périphérique, qui est flasque (ex. poliomyélite).
Il existe enfin des voies neurologiques motrices indirectes, polysynaptiques, constituant le système des voies motrices extra-pyramidales. Leur atteinte se traduit par des troubles moteurs centraux, avec rigidité de type plastique.
5.4.2.2.2.- Organisation particulière des voies motrices
5.4.2.2.2.1.- La voie motrice pyramidale : S.65

C’est une voie directe cortico-spinale. Cette voie motrice principale est responsable de la motricité volontaire. Elle descend du cortex cérébral en formant le faisceau pyramidal qui traverse la capsule interne du cerveau et se dirige verticalement vers le tronc cérébral et la moelle épinière
Dans la traversée verticale du tronc cérébral, au niveau de la moelle allongée, le faisceau pyramidal, pour sa plus grande partie (9/10 des fibres), croise la ligne médiane (décussation pyramidale) et descend dans la moelle du côté opposé à l’hémisphère cérébral d’origine. Ainsi se trouve individualisé dans le cordon latéral de la moelle, le volumineux faisceau pyramidal croisé.S.66

Un faible contingent de fibres motrices pyramidales (1/10) ne subit pas la décussation et descend dans la moelle du même côté que celui de son origine : c’est le faisceau pyramidal direct placé dans le cordon ventral, le long de la fissure médiane de la moelle.
Les axones contenus dans le faisceau pyramidal croisé se terminent du même côté en s’articulant dans la corne ventrale de la moelle avec les motoneurones alpha au niveau de chaque neuromère.
Les axones contenus dans le faisceau pyramidal direct croisent la ligne médiane au niveau de chaque neuromère et s’articulent à leur tour avec les motoneurones de la corne ventrale de la moelle du côté opposé.
En définitive, la voie pyramidale est totalement croisée. La voie motrice principale issue du cortex et descendant dans la moelle est responsable du mouvement volontaire précis et sélectif. La voie pyramidale est encore appelée voie idio-cynétique (= voie motrice particulière).
En bref, on voit que l’organisation générale de la motricité volontaire correspond à une chaîne de deux neurones, neurone central et neurone périphérique (notions de paralysie centrale et de paralysie périphérique).
5.4.2.2.2.2. - les voies motrices extra - pyramidales :
S.67

Elles sont responsables de l’activité motrice globale, consciente ou non, qui se manifeste dans le cadre du mouvement. Elles mettent en oeuvre des groupes musculaires entiers d’où leur appellation très évocatrice sur le plan fonctionnel, de voies motrices holocinétiques (= voies motrices globales). Elles proviennent des différents centres moteurs du cerveau et du tronc cérébral et parcourent verticalement les cordons de substance blanche de la moelle. Il est commode de les classer en 3 catégories : voies archéo-motrices, paléo-motrices et néo-motrices.
S.68

a) - voies extra-pyramidales archéo-motrices (les plus anciennes)
Ce sont :
– Le faisceau réticulo-spinal : issu de la substance réticulée du tronc cérébral, il comprend deux parties :
le faisceau réticulo-spinal médian à fonction facilitante. Il est placé dans le cordon ventral.
le faisceau réticulo-spinal latéral à fonction inhibitrice. Il est placé dans le cordon latéral.
– Le faisceau vestibulo-spinal : il est en rapport avec le maintien de l’équilibre. Il descend des noyaux vestibulaires de la moelle allongée et se divise en faisceau vestibulo-spinal direct et faisceau vestibulo-spinal croisé. Il est placé dans le cordon ventral.
b) - voies extra-pyramidales paléo-motrices :
Elles sont en rapport avec les mouvements globaux automatiques et inconscients. Elles comprennent :
– Le faisceau rubro-spinal : il descend du noyau rouge du tronc cérébral et plus spécialement de sa portion magno-cellulaire (appelée paléo-rubrum). Il est placé dans le cordon latéral.
– Le faisceau tecto-spinal : il descend des tubercules quadrijumeaux du tronc cérébral et il concerne les mouvements de la tête en rapport avec les sensations visuelles et auditives. Il est placé dans le cordon ventral.
c) - les voies extra-pyramidales néo-motrices :
Elles comprennent :
– Le faisceau olivo-spinal : issu de l’olive de la moelle allongée, il concerne l’activité des muscles synergiques du mouvement du membre supérieur et, de ce fait, n’existe qu’au niveau du renflement cervical de la moelle épinière. Il est placé dans le cordon ventral.
– les voies cortico-spinales extra-pyramidales : elles sont nombreuses et importantes. Elles proviennent du cortex du cerveau. Elles descendent dans la moelle, mélées aux fibres du faisceau pyramidal (Elles sont étudiées et dessinées dans le chapitre de systématisation des voies du cervelet).
5.4.2.2.2.3. - Terminaison des voies motrices
Les axones moteurs centraux des voies pyramidales et extra-pyramidales s’articulent au niveau de chaque neuromère, dans la corne ventrale avec les motoneurones périphériques. Le corps du motoneurone donne un axone moteur qui quitte la moelle par la racine ventrale des nerfs spinaux puis pénètrent dans les troncs nerveux périphériques. L’axone présente une arborisation terminale. Chaque branche apporte l’innervation motrice à une fibre musculaire striée, au niveau de la plaque motrice. Un motoneurone, son corps cellulaire, son axone et les fibres musculaires qui en dépendent constituent une UNITE MOTRICE (ou grappe neuromyonique).
Structure de la plaque motrice ou synapse neuro-musculaire :
C’est l’organe au niveau duquel l’influx nerveux arrive au contact du muscle et provoque sa contraction. Chaque branche de l’arborisation terminale du neurone s’applique sur la face latérale d’une fibre musculaire élémentaire au niveau d’un épaississement appelé plaque motrice. Au voisinage de la plaque motrice, le rameau nerveux perd sa gaine de myéline et se subdivise en plusieurs petits boutons pédiculés, appelés boutons synaptiques, qui s’implantent dans le sarcoplasme de la fibre musculaire.
Entre la paroi du bouton (membrane pré-synaptique) et la paroi de la fibre musculaire (membrane post-synaptique), il existe un petit espace appelé espace synaptique. A l’intérieur du bouton pré-synaptique, il existe des vésicules de neuro-transmetteurs qui libèrent leur contenu dans l’espace inter-synaptique. Il en résulte une dépolarisation de la membrane de la fibre musculaire qui se traduit par la contraction de la fibre.Ce mécanisme de transmission de l’influx nerveux au contact de la fibre musculaire peut être bloqué par le curare.
La myasthénie (maladie d’Erb-Goldflam) résulte d’une inefficacité de la transmission neuro-musculaire, soit par insuffisance de neurotransmetteurs (ici l’acétylcholine), soit par insensibilité de la plaque motrice à l’acétylcholine. Un certain nombre de médicaments facilitent la transmission neuro-musculaire dans ce cas (Ex. prostygmine).
5.4.3. -Moelle épinière : Fonctions réflexes S.69

Après avoir étudié les voies ascendantes (sensitives) et descendantes (motrices), il est logique d’étudier les connexions intermédiaires qui relient, dans la moelle elle-même, les structures sensitives et motrices. Elles réalisent des boucles réflexes, les unes courtes, mono ou pauci - synaptiques, les autres plus longues, poly-synaptiques, s’étendant en hauteur sur plusieurs neuromères, ou en largeur à destinée contro-latérale, par des fibres utilisant les commissures grises antérieures et postérieures de la moelle et le faisceau propre (ou fondamental).
Dans cette organisation, le faisceau propre joue un rôle important dans les relations inter-segmentaires, mais il n’est pas le seul. En effet, les fibres sensitives des cordons dorsaux ont un trajet ascendant qui formera plus haut la voie lemniscale. Mais, dans chaque neuromère, ces fibres donnent des collatérales descendantes plus courtes, qui assurent des relations d’association intersegmentaire, sur quelques neuromères, avec prolongement contro-latéral. Ces groupes de fibres descendantes sont décrites, dans les cordons dorsaux sous le nom de faisceaux de Schultze, de Hoche et de Gombault. De plus, toutes les fibres sensitives envoient des collatérales sur les interneurones de la lame VII.
Globalement, nous sommes, ici, en présence d’un important système associatif intra-médullaire, à fonctionnement réflexe. Ainsi s’explique la diffusion uni ou bilatérale de l’action réflexe, en fonction de l’intensité de la stimulation. Ce fonctionnement réflexe est normalement régulé par l’action modulatrice des centres supérieurs (action inhibitrice de la voie cortico-réticulo-spinale dont l’interruption pathologique (Ex : paraplégie) aboutit à la spasticité.
A - Appareil de connexion intra-médullaire (Faisceau propre ou faisceau fondamental).
– en hauteur : la diffusion pluri-segmentaire est assurée par le faisceau propre (ou fondamental) ainsi que par les collatérales descendantes des fibres sensitives intra-médullaires des systèmes lemniscal, extra-lemniscal et spino-cérébelleux ; L’ensemble de ces collatérales s’articule dans la lame VII (zone des interneurones) avec les cellules d’association du faisceau fondamental.
– en largeur : la structure de la lame VII contient des fibres commissurales assurant la diffusion controlatérale.
B - Réflexes normaux ou spontanés
Deux catégories : réflexes courts, réflexes longs.
1 - Réflexes courts : monosynaptiques ou pauci - synaptiques Fibres nerveuses motrices et boucle gamma). S.70
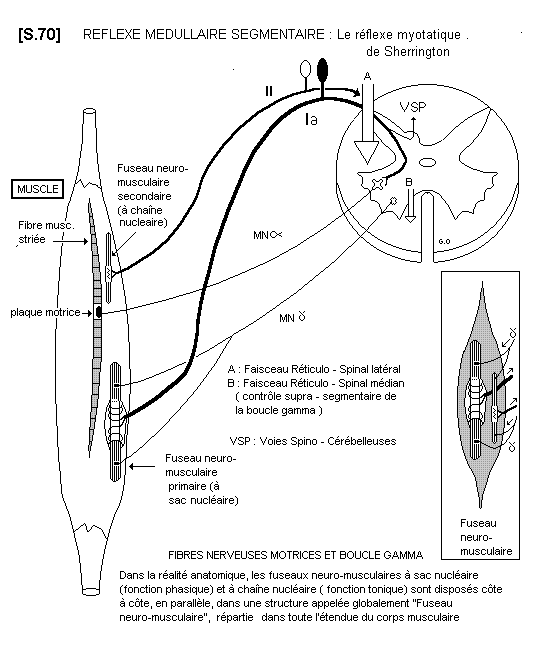
Le prototype de ce système est le réflexe myotatique de la boucle gamma. C’est un réflexe proprioceptif inconscient. Les sources sensitives intramusculaires du réflexe sont les fuseaux neuro - musculaires : fuseaux primaires (phasiques) pour le réglage de la longueur des fibres musculaires, et fuseaux secondaires (toniques) pour le réglage de la tension des fibres musculaires. Les organes neuro - tendineux de Golgi sont aussi des capteurs de tension réglés à un seuil plus élevé que les précédents, pour la protection de la jonction musculo - tendineuse.
La régulation des paramètres musculaires dans le mouvement normal est assurée par l’action modulatrice de la voie cortico - réticulo - spinale, citée ci-dessus. La pathologie motrice centrale démontre la puissance de ce mécanisme réflexe, quand il n’est plus régulé (= spaticité).
2- Réflexes longs : polysynaptiques : plus étendus et plus complexes, ils sont présents dans plusieurs comportements moteurs et variables en fonction du degré de maturation neurologique.
Chez le nouveau né (1er mois), les réflexes sont appelés par les pédiatres : automatismes primaires, en raison de l’immaturité de la myélinisation de la voie pyramidale et des connexions synaptiques. Ce sont :
– réflexe de succion (déjà présent avant la naissance). C’est un réflexe du tronc cérébral, mais cité ici en raison de son importance chez le nouveau-né.
– réflexe de Moro : Extension/abduction des 4 membres, à la dépression du lit (disparaît au 3ème mois)
– réflexe tonique du cou : flexion des membres du côté de la tête, extension de l’autre
– réflexe d’allongement croisé du MI, à la stimulation plantaire opposée
– « grasping » (empoignement)
Cette période d’hyper - réflexie est appelée : « Stade pré-pyramidal ».
Chez le nourrisson (de 2 à 18 mois) :
– Fin de l’hypertonie de flexion.
– Présence des réflexes rotulien et achilléen, mais diffusés
– Réflexes de redressement se renforçant progressivement : Tient la tête à 3 mois.
– réflexe d’incurvation dorsale du tronc (en position ventrale)
– Déplacements à plat ventre dans le lit (« locomotion ichtyoïde » et réflexes de flexion)
– Appui sur les coudes (« locomotion reptilienne et réflexes d’extension vers 5 mois
– réflexe de marche automatique et d’enjambement
– Position assise et déplacement à quatre pattes vers 8 mois
– Verticalisation et bipédie avec appui vers 10 - 12 mois
– La marche est acquise à 15 mois mais solide à 18 mois
Il y a donc une évolution de l’activité réflexe, aidée par l’apprentissage.
Chez l’adulte, les réflexes spontanés sont principalement extéroceptifs, nociceptifs et proprioceptifs :
– Aux membres supérieurs : Réflexes de flexion (ou de protection ou de fuite).Ils diffusent par la voie de neurones intersegmentaires (Ex : toucher un objet brûlant : retrait d’un ou deux membres.
– Aux membres inférieurs : appui douloureux de la plante du pied : retrait en triple flexion avec réflexe croisé d’extension opposée, pour assurer l’appui au sol.Mouvements de rattrapage d’équilibre
C - Réflexes provoqués :
Les réflexes provoqués sont largement utilisés en séméiologie. Ils sont à point de départ ostéo - tendineux ( proprioceptifs) ou cutanés (extéroceptifs ). Ils étudient, à la fois, l’état des voies périphériques du réflexe (n. sensitif / n. moteur) et l’état de la réflectivité centrale.
Les réflexes en séméiologie : Tableau des réflexes
TABLEAU DES PRINCIPAUX REFLEXES
TABLEAU

A noter que les cinq premiers sont des réflexes tendineux et les quatre derniers, des réflexes cutanés
D - Synthèses
– 1-Neuro - physiologie et Théorie de l’information
La théorie de l’information s’applique bien aux fonctions du Système nerveux central, et spécialement aux fonctionnements réflexes.
Exemple : Source de l’information sensitive ou sensorielle => transfert (avec relais et diffusion éventuels) =>destination, analyse et traitement => départ de l’information motrice => voies motrices directes ou indirectes => réponse réflexe motrice. Cette transmission comporte des contrôles en feed-back.
La physio - pathologie de la fonction motrice se situe nécessairement dans l’une de ces étapes, soit au niveau des voies et des organes nerveux proprement dits, soit au niveau des champs synaptiques qui les unissent.
– 2-Techniques de facilitation et d’inhibition en rééducation fonctionnelle
Ce sont des méthodes globales recrutant des chaînes musculaires cinétiques et des groupements musculaires antagonistes.
La diffusion ipsi et contro - latérale des réflexes médullaires, exposée ci-dessus est un argument neuro - physiologique à mettre au crédit des méthodes de rééducation neuromusculaire proprioceptive.( Méthodes de Kabat, de Knot, de Bobath et de Rood (Reprogrammation neuro - motrice de Viel) . ( réf. Moody J.M., Noël F. et Viel E. - Méthodes de rééducation neuromusculaire - Encycl. Méd. Chir. Paris - Kinésithérapie, 3-23-01, 26060 A10 ).
Il faut signaler aussi, pour son importance, ses résultats et sa relative simplicité, la rééducation musculaire à base de réflexes posturaux de Gilbert (réf. Gilbert M.A. et Adam M.- Encycl.Méd. Chir. Paris - Kinésithérapie - 4-5-09 - 26061 - A10.
– 3- Réflexothérapie ou massage réflexe (Bindegewebs massage)
5.4.4.- Coupe horizontale théorique de la moelle épinière : S.71

Tous les faisceaux nerveux existant dans la substance blanche de la moelle épinière sont représentés sur un dessin de synthèse de coupe de la moelle. Il s’agit d’une coupe théorique, puisque tous les faisceaux ne sont pas présents à tous les niveaux de la moelle (les faisceaux qui concernent le membre supérieur ne sont présents que dans le renflement cervical).
5.4.5.- Applications cliniques (Les principaux syndromes médullaires)
– a) Paraplégie spasmodique : Déficit moteur des deux membres inférieurs avec spasticité. (Compression de moelle ou section de moelle).
– b) Syndrome de Brown - Sequard (Section de l’hémi - moelle).
Dans le territoire sous-jacent :
Hémiplégie avec spasticité, du côté de la lésion
Abolition des sensibilités extra-lemniscales (thermo-algésiques) du côté opposé à la lésion
Atteinte (incomplète) des sensibilités spino-cérébelleuses du côté de la lésion
Atteinte, plus ou moins complète, des sensibilités lemniscales (tact épicritique)
du côté de la lésion.
Il s’agit d’un syndrome rarement pur.
A retenir : l’association d’un syndrome pyramidal d’un côté, et d’un syndrome spino-thalamique de l’autre, est caractéristique de l’hémi-lésion médullaire, et a une grande valeur localisatrice. En bref, il s’agit d’un syndrome alterne de la moelle.
– c) Syndrome de la corne ventrale : Exemple : Poliomyélite
– d) Syndrome des cordons dorsaux : Atteinte des sensibilités lemniscales, et spécialement de la sensibilité proprioceptive consciente (sens articulaire ou sensibilité profonde). (Notion d’ataxie radiculo - cordonnale = Tabès).
– e) Syndrome syringomyélique : dissociation des sensibilités
– f) Syndrome de la sclérose combinée de la moelle : Atteinte pyramidale + Syndrome des cordons postérieurs de la moelle.
Pr Gérard Outrequin - Dr Bertrand Boutillier - Toute reproduction interdite
Licence d'utilisation - Mentions légales - Politique de confidentialité - Faire un don - Découvrez MedShake !